3. Big functions from small circuits¶
Design principles
Increased protein turnover speeds up the response time of a gene expression system.
Negative autoregulation accelerates turn-on times.
Positive, ultrasensitive feedback enables bistability.
Key concepts
Demand theory relates the fraction of time a gene is expressed to its mode of regulation.
Decay rates determine response times
Techniques
Steady state normalization allows analysis of response times.
Nullclines identify fixed points and provide insight into the structure of dynamical systems.
[1]:
# Colab setup ------------------
import os, sys, subprocess
if "google.colab" in sys.modules:
cmd = "pip install --upgrade watermark"
process = subprocess.Popen(cmd.split(), stdout=subprocess.PIPE, stderr=subprocess.PIPE)
stdout, stderr = process.communicate()
# ------------------------------
import numpy as np
import scipy.integrate
import scipy.optimize
import bokeh.io
import bokeh.models
import bokeh.plotting
bokeh.io.output_notebook()
Demand theory: “use it or lose it”¶
At the end of the first chapter, we asked our first design question: Under what conditions would you prefer to use activation or repression to regulate a gene? Both schemes are used in nature. They can each generate equivalent responses–a signal or condition that activates an activator should behave similarly to a signal that inhibits a repressor:
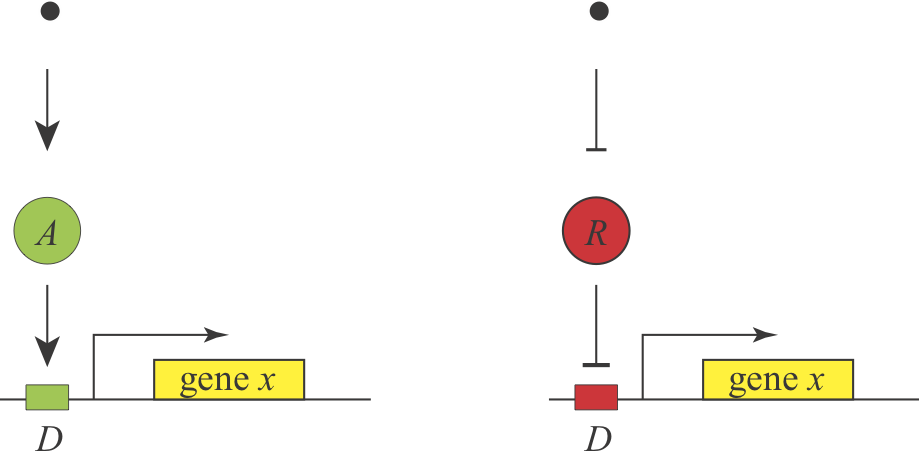
Michael Savageau posed this question in the context of bacterial metabolic gene regulation in his paper, “Design of molecular control mechanisms and the demand for gene expression” (PNAS, 1977). He focused on “demand” as a critical factor that influences the choice of activation or repression. Demand can be defined as the fraction of time that the gene is needed at the high end of its regulatory range in the cells natural environments. Savageau made the empirical observation that high demand genes are more frequently regulated by activators, while low demand genes are more often regulated by repressors.
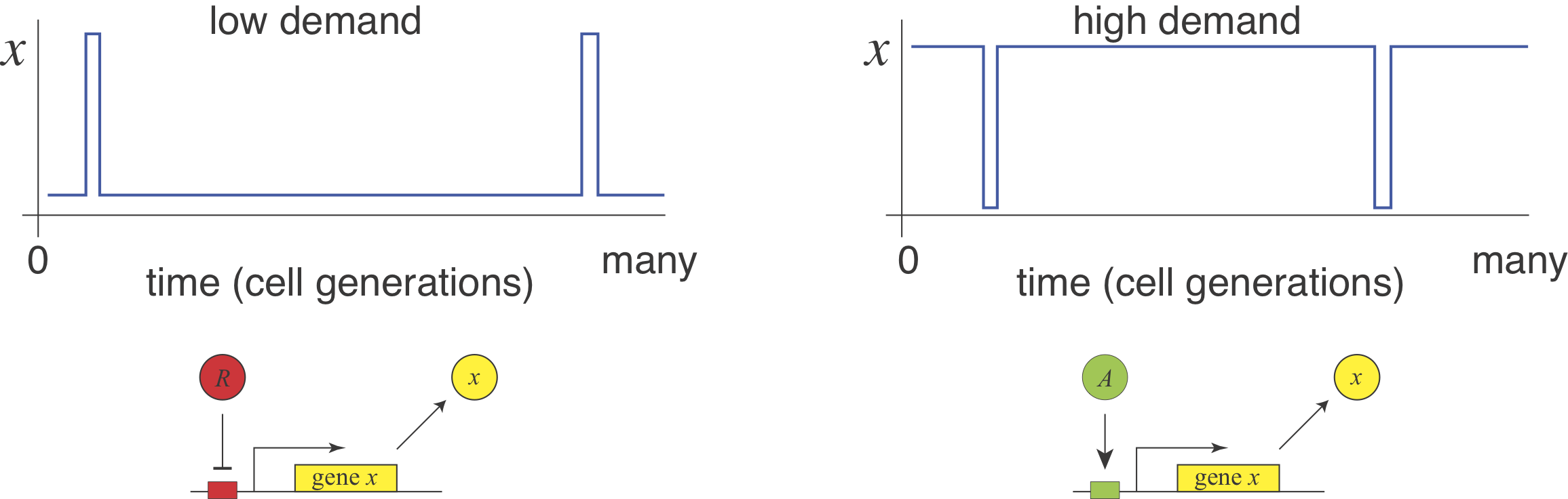
He suggested that this relationship could be explained by a “use it or lose it” rule of evolutionary selection. Recall that mutations, especially those that diminish the function of a gene, occur frequently. A high demand gene controlled by an activator needs the activator to be on most of the time. Under these conditions, evolution would select against mutations that eliminate the activator. By contrast, if the same high demand system were regulated by a repressor then, most of the time, there would be weaker selection against mutations that removed the repressor, increasing the potential for evolutionary loss of the regulatory system. This reasoning assumes no direct fitness advantage for either regulation mode, just a difference in the average strength of selection pressure needed to maintain them.
In 2009, Gerland and Hwa formulated and analyzed a model to explore these ideas mathematically. They showed that the “use it or lose it” principle indeed dominates when timescales of switching between low and high demand environments are long and populations are small. However, the same model could select for the opposite demand rule in other regimes.
Shinar, et al. introduced a different explanation for the demand rules. The authors assumed that “naked” DNA binding sites, which are not bound to proteins, are susceptible to non-specific binding of transcription factors. These inadvertent encounters between transcription factors and DNA could lead to inappropriate activation or repression of adjacent genes, imposing a low, but non-zero fitness cost. “Intentionally” keeping these sites occupied most of the time minimizes the fraction of time that such events can occur.
This explanation make the same predictions as Savageau’s explanation but for different reasons. Here, a high demand gene should preferentially use an activator since this arrangement minimizes unoccupied binding sites more of the time. Conversely, a low demand gene would preferentially use repression to maintain the binding site in an occupied state under most conditions. This argument can be generalized to other examples of seemingly equivalent regulatory systems and is described in Uri Alon’s book.
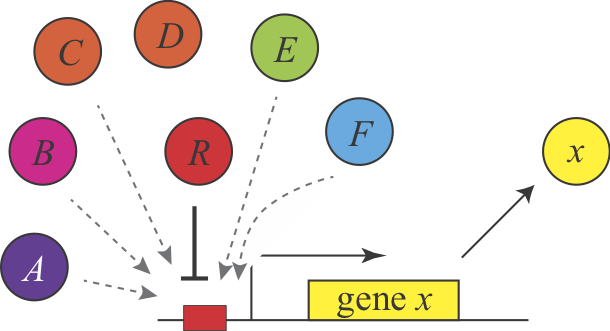
In this figure, a repressor, denoted R binds strongly and specifically to its target site. In the absence o the repressor, that site could also be bound non-specifically, and inappropriately by a range of other factors, denoted A-F.
Remarkably, we still lack definitive experimental evidence to fully resolve this fundamental design question. Challenge: Can you devise an experimental way to discriminate among these potential explanations?
Dynamics: protein stability determines the response time to a change in gene expression¶
We now return to simple gene regulation circuits. So far, we have focused on steady states. However, many biological circuits change dynamically, even in constant environmental conditions.
The most basic dynamic question one can ask about a gene is how rapidly it can switch from one level of protein to a new level after a sudden increase or decrease in gene expression.
Starting with our basic equation for gene expression, and neglecting the mRNA level for the moment, we write:
\begin{align} \frac{\mathrm{d}x}{\mathrm{d}t} = \beta - \gamma x \end{align}
Consider a situation in which the gene has been off for a long time, so that \(x(t=0)=0\) and then is suddenly turned on at \(t=0\). Its dynamics then follow,
\begin{align} x(t) = \frac{\beta }{\gamma} (1-e^{-\gamma t}) \end{align}
Thus, the parameter that determines the response time is \(\gamma\), the degradation+dilution rate of the protein. We define the response time (indicated by the vertical dashed line on the plot below) to be \(\gamma^{-1}\), the time it takes for the concentration to rise to a factor of \(1-\mathrm{e}^{-1}\) of its steady state value.
[2]:
# Parameters
beta = 100
gamma = 1
# Dynamics
t = np.linspace(0, 6, 400)
x = beta / gamma * (1 - np.exp(-gamma * t))
# Plot response
p = bokeh.plotting.figure(
frame_height=275,
frame_width=375,
x_axis_label="t",
y_axis_label="x(t)",
title=f"β = {beta}, γ = {gamma}",
x_range=[0, 6],
)
p.line(t, x, line_width=2)
# Mark the response time (when we get to level 1-1/e)
t0 = 1 / gamma
x0 = beta / gamma * (1 - np.exp(-1))
# Add the glyph with label
source = bokeh.models.ColumnDataSource(
data=dict(t0=[t0], x0=[x0], text=["response time"])
)
p.circle(x="t0", y="x0", source=source, size=10)
p.add_layout(
bokeh.models.LabelSet(
x="t0", y="x0", text="text", source=source, x_offset=10, y_offset=-10
)
)
p.add_layout(
bokeh.models.Span(
location=t0,
level="underlay",
dimension="height",
line_color="black",
line_dash="dashed",
)
)
bokeh.io.show(p)
How can we speed up responses?¶
So far, it seems as if the cell’s ability to modulate the concentration of a stable protein is quite limited, apparently requiring multiple cell cycles for both increases and decreases. This seems rather pathetic for such a successful, billion year old creature. You might think you could up-regulate the protein level faster by cranking up the promoter strength (increasing \(\beta\)). Indeed, this would allow the cell to hit a specific threshold earlier. However, it would also increase the final steady state level (\(\beta/\gamma\)), and therefore leave the timescale over which the system reaches its new steady-state unaffected.
One simple and direct way to speed up the response time of the protein is to destabilize it, increasing \(\gamma\). This strategy pays the cost of a “futile cycle” of protein synthesis and degradation to provide a benefit in terms of the speed with which the regulatory system can reach a new steady state.
To disentangle amplitude and timescale, it is useful to normalize these plots by their steady states, as in the right-hand plot below. Comparing normalized response curves is reasonable because many mutations can alter the expression level of a gene allowing evolution the potential to optimize expression level independently of the regulatory feedback system.
[3]:
# Parameters
beta_1 = 100
gamma = np.array([1, 2, 3])
# Compute dynamics
t = np.linspace(0, 6, 400)
x = [beta_1 / g * (1 - np.exp(-g * t)) for g in gamma]
# Set up plots
colors = bokeh.palettes.Oranges5
p1 = bokeh.plotting.figure(
frame_height=175,
frame_width=300,
x_axis_label="t",
y_axis_label="x(t)",
title=f"β₁ = {beta_1}, γ = {gamma}",
x_range=[0, 6],
)
p2 = bokeh.plotting.figure(
frame_height=175,
frame_width=300,
x_axis_label="t",
y_axis_label="x(t)",
title="same plot, normalized by steady states",
x_range=[0, 6],
)
p2.x_range = p1.x_range
# Populate graphs
for x_vals, g, color in zip(x, gamma, colors):
p1.line(t, x_vals, color=color, line_width=2)
p1.circle(1 / g, beta_1 / g * (1 - np.exp(-1)), color=color, size=10)
p2.line(t, x_vals / x_vals.max(), color=color, line_width=2)
p2.circle(1 / g, 1 - np.exp(-1), color=color, size=10)
# Label lines
p1.text(
x=[4],
y=[95],
text=[f"γ = {gamma[0]}"],
text_color=colors[0],
text_font_size="10pt",
text_align="left",
text_baseline="top",
)
p1.text(
x=[4],
y=[53],
text=[f"γ = {gamma[1]}"],
text_color=colors[1],
text_font_size="10pt",
text_align="left",
text_baseline="bottom",
)
p1.text(
x=[4],
y=[30],
text=[f"γ = {gamma[2]}"],
text_color=colors[2],
text_font_size="10pt",
text_align="left",
text_baseline="top",
)
bokeh.io.show(bokeh.layouts.row(p1, bokeh.models.Spacer(width=20), p2))
Design principle: Increased turnover speeds up the response time of a gene expression system, at the cost of additional protein synthesis and degradation.
Negative autoregulation is prevalent in natural circuits¶
So destabilizing a protein can speed its resposne time. However, most bacterial proteins, transcription factors in particular, are stable. Do they use other mechanisms to accelerate response times?
Empirically, many repressors repress their own expression.
In fact, we can consider autoregulation to be a network motif, defined as a regulatory pattern that is statistically over-represented in natural networks (circuits) compared to random networks. To see this, it is useful to imagine the transcriptional regulatory network of an organism as a graph consisting of nodes and directed edges (arrows). In bacteria, each node represents an operon, while each arrow represents regulation of the target operon (tip of the arrow) by a transcription factor in the originating operon (base of the arrow), as shown schematically here.
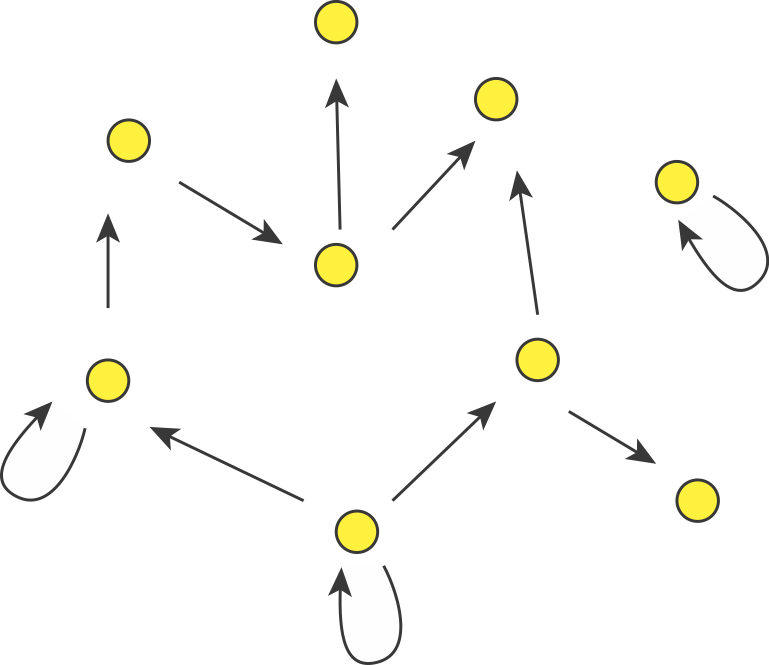
The transcriptional regulatory network of E. coli has been mapped (see RegulonDB). It contains ≈424 operons (nodes), ≈519 transcriptional regulatory interactions (arrows), involving ≈116 transcription factors. If the target of each arrow was chosen randomly, the probability of any given arrow being autoregulatory is low (≈1/424). One might expect only about one such event in the entire network. However, ≈40 such autoregulatory arrows are observed. Autoregulation thus appears to be over-represented.
(If we further consider the “sign” of the arrow, with “+” representing activation and “-” representing repression, it turns out that there are 32 negative autoregulatory operons and 8 positive autoregulatory ones. We will discuss both types.)
This is a simple example of the motif principle, which states that statistically over-represented patterns in networks have been selected for key cellular functions. In fact, the motif concept is more general. For example, sequence motifs are statistically over-represented sequences within the genome that are enriched for functionally important features, such as protein binding sites. This is a core concept in bioinformatics. We will discuss network motifs further in the next chapter.
Negative autoregulation accelerates response times¶
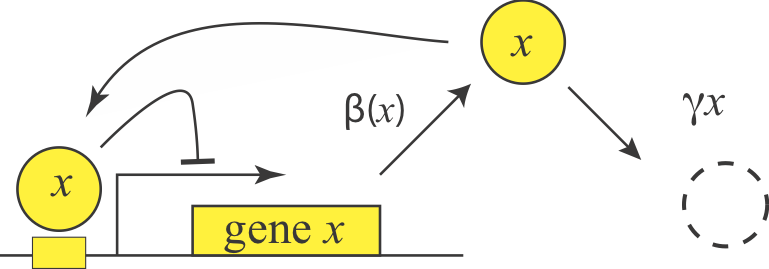
Now that we know negative autoregulation is prevalent, we ask what function or functions it provides for the cell. To start, we write down a differential equation representing production and degradation of the repressor, \(x\). We can incorporate repression using the expression we derived in the first chapter, except here, the repressor is \(x\) itself.
\begin{align} \frac{\mathrm{d}x}{\mathrm{d}t}=\frac{\beta}{1+x/K_d} - \gamma x \end{align}
We will consider the limit in which the autoregulation is “strong”, i.e. where \(\beta/\gamma \gg K_d\), so the gene can, at maximal expression level, produce enough protein to fully repress itself.
What happens when the operon is suddenly turned “on” from an initial “off” state (\(x(0)=0\))? Initially, \(x\) builds up approximately linearly, at rate \(\beta\). Eventually, its concentration is high enough to shut its own production off, \(x \sim K_d\). While the real dynamcs are smoother, one can think of the behavior roughly like this:
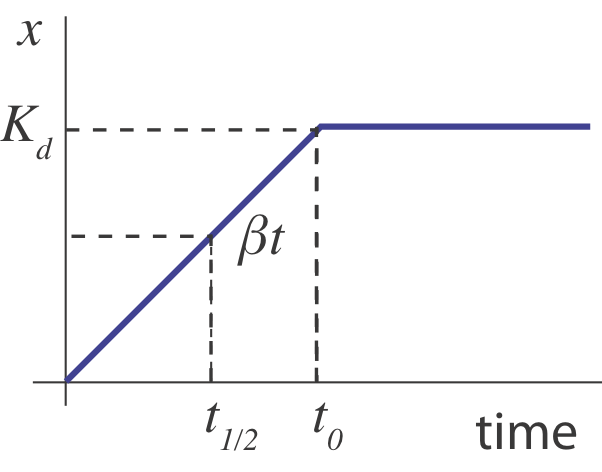
In this sketch, we can see that the half-time, \(t_{1/2}\) for turning on should occur when \(\beta t \approx K_d / 2\), i.e. at \(t \approx K_d/2 \beta\). But this is only an approximation. A more complete treatment, in Rosenfeld et al, JMB 2002 shows that in the limit of strong negative autoregulation the dynamics approach \(x(t) = x_\mathrm{st} \sqrt{1-\mathrm{e}^{-2 \gamma t}}\), where \(x_\mathrm{st}\) denotes the steady-state expression level. In the following section, we will explore these dynamics using numerical integration and compare them to this analytical approximation.
[4]:
# Negative autoregulation right hand side
def neg_auto_reg_rhs(x, t, beta, Kd, gamma):
return beta / (1 + x / Kd) - gamma * x
# Parameters
t = np.linspace(0, 6, 400)
Kd = 1
gamma = 1
beta = 100
# Negative autoregulated solution
y = scipy.integrate.odeint(neg_auto_reg_rhs, 0, t, args=(beta, Kd, gamma))
nar = y[:, 0]
# Unregulated solution
unreg = 1 - np.exp(-gamma * t)
# Limiting solution
limiting = np.sqrt(1 - np.exp(-2 * gamma * t))
# Create plots of unnormalized responses
p = [
bokeh.plotting.figure(
frame_height=250,
width=350,
x_axis_label="t",
y_axis_label="x(t)",
x_range=[0, 6],
)
for _ in [0, 1]
]
p[0].x_range = p[1].x_range
p[0].title.text = "Turn-on dynamics ± negative autoregulation"
p[0].line(
t,
nar.max() * limiting,
line_width=4,
color="orange",
legend_label="limiting analytical solution",
)
p[0].line(t, nar, line_width=2, legend_label="negative autoregulation")
p[0].line(
t, beta / gamma * unreg, line_width=2, color="grey", legend_label="unregulated",
)
p[0].legend.location = "center_right"
# Create plot of normalized responses
p[1].title.text = "Same plot, normalized"
p[1].line(t, limiting, line_width=4, color="orange")
p[1].line(t, nar / nar.max(), line_width=2)
p[1].line(t, unreg, line_width=2, color="grey")
bokeh.io.show(bokeh.layouts.gridplot(p, ncols=1))
What do we see here? First, as you would expect, adding negative autoregulation reduces the steady-state expression level. However, it also has a second effect: accelerating the approach to steady state. The second plot includes the limiting analytical solution from Rosenfeld et al. for comparison.
In fact, negative autoregulation has accelerated the dynamics by about 5-fold compared to the unregulated system.
Note that this acceleration occurs when we turn the gene on, but not when we turn it off. If we suddenly stop expression by setting \(\beta\) to 0, then the dynamics are governed by \(\mathrm{d}x/\mathrm{d}t = -\gamma x\) irrespective of which architecture we use.
Can this acceleration be observed experimentally? To find out, Rosenfeld et al. engineered a simple synthetic system based on a bacterial repressor called TetR, fused to a fluorescent protein for readout, and studied its turn-on dynamics in bacterial populations.
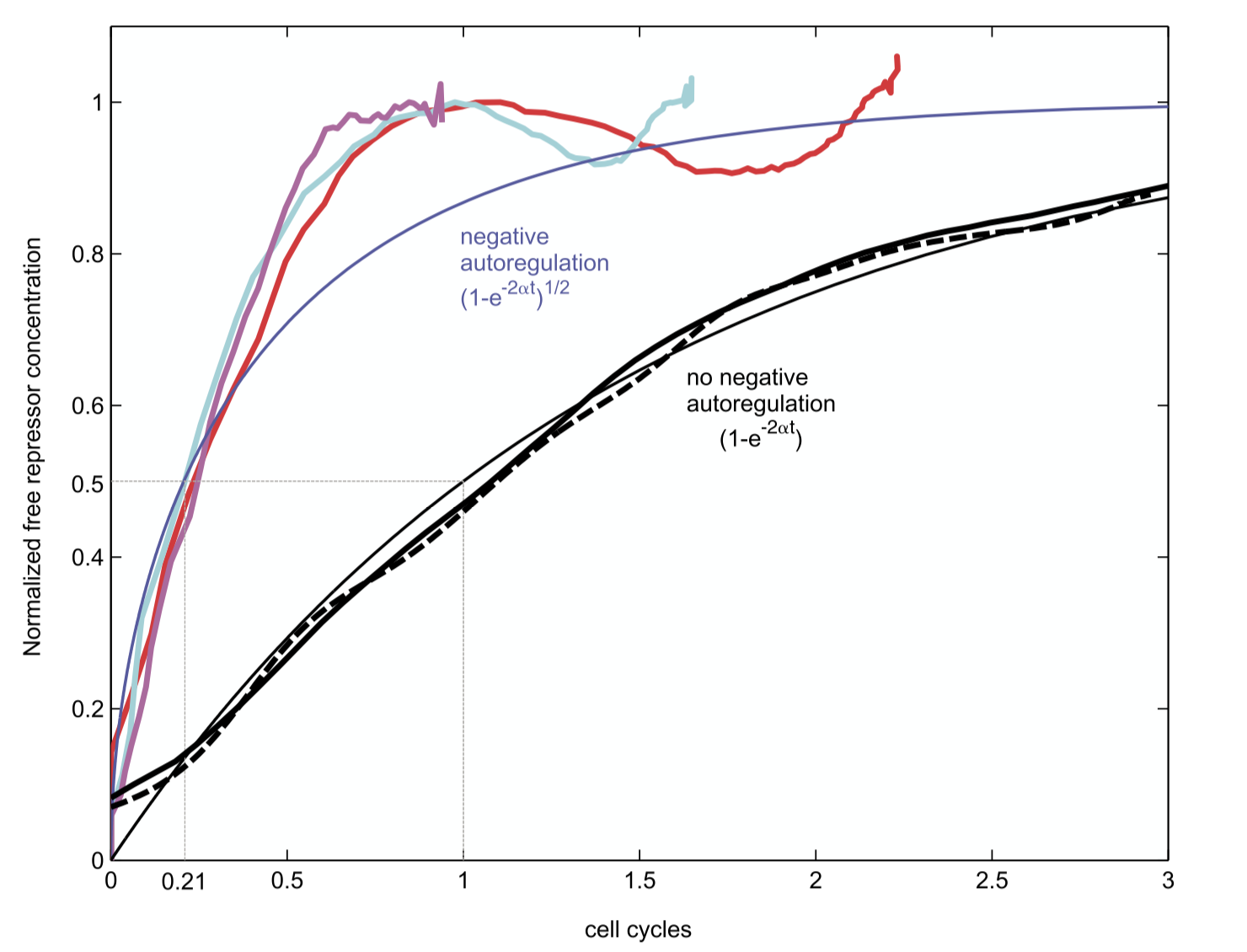
This image is taken from Rosenfeld et al., J. Mol. Biol., 2002.
Interestingly, these dynamics showed the expected acceleration, as well as some oscillations around steady-state, which may be explained by time delays in the regulatory system (something we will discuss more in an upcoming chapter).
To conclude this section: We now have identified another simple design principle: Negative autoregulation speeds the response time of a transcription factor.
Negative autoregulation can have additional functions beyond acceleration. Using similar synthetic approaches, negative autoregulation was shown to reduce stochastic cell-cell variability (“noise”) in gene expression (Becskei and Serrano, Nature, 2000).
Interlude: Hill functions and ultrasensitivity¶
Before moving forward, we will take a moment to introduce the Hill function, which we briefly visited in the previous chapter. Many responses in gene regulation and protein-protein interactions have a switch-like, or ultrasensitive shape. Ultrasensitivity can arise from cooperativity in molecular interactions. For example, consider a situation in which binding of a protein at one DNA binding site increases the affinity for binding of a second protein at an adjacent site. Or, imagine a protein with an alternative molecular conformation that is stabilized by binding of multiple agonist effector molecules and, in that conformation, has a higher affinity for the same effectors. In these and many other situations, an increasing concentration of one species can have little effect for a while, and then suddenly have a large effect.
The Hill function provides a way to analyze systems that exhibit ultrasensitive responses. While it can be derived from models of some processes, it is often used in a more generic way to analyze how a circuit would behave with different levels of ultrasensitivity.
An activating Hill function is defined by
\begin{align} f_\mathrm{act}(x) &= \frac{x^n}{k^n +x^n} = \frac{(x/k)^n}{1 + (x/k)^n}. \end{align}
You can also make a mirror image repressive Hill function.
\begin{align} f_\mathrm{rep}(x) &= \frac{k^n}{k^n +x^n} = \frac{1}{1 + (x/k)^n}. \end{align}
In these expressions, \(k\) is often referred to as an activation coefficient; it represents the concentration at which the function attains half its maximal value. It is a measure of the concentration \(x\) that is necessary to affect the regulation. The Hill coefficient, \(n\), parametrizes how ultrasensitive the response is. When \(n=1\), we recover the simple binding curves introduced earlier. When \(n>\), however, we achieve ever sharper, more ultrasensitive, responses. In the limit of \(n=\infty\) we have a step function. Here we plot the Hill function for a few values of \(n\):
[5]:
x = np.logspace(-2, 2, 200)
f = [x**n / (1 + x**n) for n in [1, 2, 10]]
# Build plot
p = bokeh.plotting.figure(
height=275,
width=400,
x_axis_label="x/K",
y_axis_label="activating Hill function",
x_range=[x[0], x[-1]],
x_axis_type="log"
)
colors = bokeh.palettes.Blues5[1:-1][::-1]
for f_act, n, color in zip(f, [1, 2, 10], colors):
p.line(x, f_act, line_width=2, color=color, legend_label=f"n = {n}")
p.legend.location = "center_right"
bokeh.io.show(p)
Try altering the code above to analyze repressive Hill functions (or play with sliders as in the previous chapter). What happens when you try negative Hill coefficients?
The name of the Hill function is suggestive–it looks like a hill. In fact, it was named for Archibald Hill, who used it to model binding of oxygen by Hemoglobin.
Positive autoregulation enables bistability¶
Having examined negative autoregulation, we now turn to its opposite, positive autoregulation, a feature that is also prevalent in natural circuits. What functions could positive autoregulation provide?
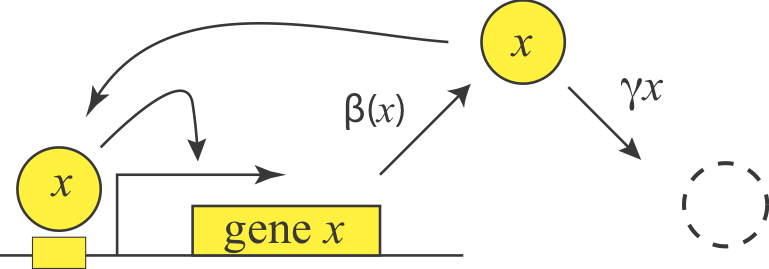
We can represent a positive autoregulatory circuit with the following simple equation, where the production term comes from our analysis of an activation system in chapter 1:
\begin{align} \frac{\mathrm{d}x}{\mathrm{d}t} = \frac{\beta x^n} {x^n + K^n} - \gamma x \end{align}
Note that we’ve replaced the simple activation function with a more general Hill function. To see what positive autoregulation might do, let’s plot the two terms on the right hand side (production rate and removal rate) versus \(x\). We will start by considering a relatively high Hill coefficient of \(n=4\).
[6]:
# Parameters
beta = 10
gamma = 1
K = 4
n = 4
# Theroetical curves
x = np.linspace(0, 20, 400)
prod = beta * x ** n / (x ** n + K ** n)
removal = gamma * x
# Fixed points
fp_rhs = lambda x: beta / gamma * x ** n / (x ** n + K ** n)
fixed_points = [
0,
float(scipy.optimize.fixed_point(fp_rhs, 3)),
float(scipy.optimize.fixed_point(fp_rhs, 5)),
]
# Build plot
p = bokeh.plotting.figure(
frame_height=275,
frame_width=375,
x_axis_label="x",
y_axis_label="production or removal rate",
title=f"β = {beta}, γ = {gamma} , K = {K}, n = {n}",
x_range=[-1, 20],
)
# Plot production and removal rates
p.line(x, prod, line_width=2, color="#1f77b4")
p.line(x, removal, line_width=2, color="orange")
# Plot fixed points
for i, fp in enumerate(fixed_points):
p.add_layout(
bokeh.models.Span(
location=fp,
level="underlay",
dimension="height",
line_color="black",
line_dash="dashed",
)
)
fill_color = "white" if i % 2 else "black"
p.circle(
[fp],
[gamma * fp],
color="black",
size=10,
line_width=2,
fill_color=fill_color,
)
# Annotate
p.add_layout(
bokeh.models.Arrow(
end=bokeh.models.VeeHead(
size=15, fill_color="gray", line_color="gray"
),
line_width=4,
x_start=3.25,
y_start=12,
x_end=0.1,
y_end=12,
line_color="gray",
)
)
p.add_layout(
bokeh.models.Arrow(
end=bokeh.models.VeeHead(
size=15, fill_color="gray", line_color="gray"
),
line_width=4,
x_start=3.5,
y_start=12,
x_end=6.75,
y_end=12,
line_color="gray",
)
)
p.add_layout(
bokeh.models.Arrow(
end=bokeh.models.VeeHead(
size=15, fill_color="gray", line_color="gray"
),
line_width=4,
x_start=20,
y_start=12,
x_end=13,
y_end=12,
line_color="gray",
)
)
p.text(
x=[14],
y=[9.933],
text=["production rate"],
text_color="#1f77b4",
text_font_size="10pt",
text_align="left",
text_baseline="top",
)
p.text(
x=[14],
y=[14],
text=["removal rate"],
text_color="orange",
text_font_size="10pt",
text_align="left",
angle=0.6,
)
bokeh.io.show(p)
Wherever production rate = degradation rate, we have a fixed point. For these parameters, there are three fixed points. These points differ in their stability. The white one in the middle is unstable, while the two black ones on the ends are stable. The easiest way to see this is to notice that between the first and second fixed points removal rate > production rate, and hence x will decrease, while between the second and third fixed points, production exceeds removal, and x will increase.
Since this system has two stable fixed points, we can describe it as bistable. As long as noise or other perturbations are not too strong, the cell can happily remain at either a low or high value of x. Bistability is a special case of the more general phenomenon of multistability, which is one of the most important properties in biology, underlying the ability of a single genome to produce a vast array of distinct cell types in a multicellular organism. This simple analysis shows us immediately that a single positive autoregulatory gene can generate bistability!
However, positive feedback by itself is not enough. We also need an ultrasensitive response to \(x\). (In the context of the Hill function, ultrasensitivity can be defined as \(n>1\)). If we reduce the Hill coefficient to 1, keeping other parameters the same, you can see that we now have only a single stable fixed point (and one unstable fixed point at \(x=0\)).
[7]:
# Reduce n to 1
n = 1
# Recompute production curve
prod = beta * x ** n / (x ** n + K ** n)
# Fixed points
fp_rhs = lambda x: beta / gamma * x ** n / (x ** n + K ** n)
fixed_points = [0, float(scipy.optimize.fixed_point(fp_rhs, 3))]
# Build plot
p = bokeh.plotting.figure(
frame_height=275,
frame_width=375,
x_axis_label="x",
y_axis_label="production or removal rate",
title=f"β = {beta}, γ = {gamma} , K = {K}, n = {n}",
x_range=[-1, 20],
)
# Plot production and removal rates
p.line(x, prod, line_width=2, color="#1f77b4")
p.line(x, removal, line_width=2, color="orange")
# Plot fixed points
for i, fp in enumerate(fixed_points):
fill_color = "black" if i % 2 else "white"
p.circle(
[fp],
[gamma * fp],
color="black",
size=10,
line_width=2,
fill_color=fill_color,
)
bokeh.io.show(p)
Furthermore, ultrasensitivity is also not enough. Bistability in this system further requires tuning of different rate constants. For example, consider what happens for varying values of \(\gamma\). (Here, I’ve set a very high value of \(n=10\) just to focus on the role of \(\gamma\).)
[8]:
n = 10 # highly cooperative
gamma = [1, 2]
# Recompute production rate
prod = beta * x ** n / (x ** n + K ** n)
# Fixed points
fp_rhs = lambda x: beta / gamma[0] * x ** n / (x ** n + K ** n)
fixed_points = [
0,
float(scipy.optimize.fixed_point(fp_rhs, 4)),
float(scipy.optimize.fixed_point(fp_rhs, 10)),
]
# Build plot
p = bokeh.plotting.figure(
frame_height=275,
frame_width=375,
x_axis_label="x",
y_axis_label="production or removal rate",
title=f"β = {beta}, γ = {gamma} , K = {K}, n = {n}",
y_range=[-1, 21],
x_range=[-1, 20],
)
# Plot production and removal rates
p.line(x, prod, line_width=2, color="#1f77b4")
p.line(x, gamma[0] * x, line_width=2, color="orange")
p.line(x, gamma[1] * x, line_width=2, color="orange")
# Plot fixed points
for i, fp in enumerate(fixed_points):
fill_color = "white" if i % 2 else "black"
p.circle(
[fp],
[gamma[0] * fp],
color="black",
size=10,
line_width=2,
fill_color=fill_color,
)
bokeh.io.show(p)
Note that with just a two-fold higher value of \(\gamma=2\) (upper orange line), we lose the last two fixed points, giving us a monostable system that just remains, sadly, at 0, totally unable to activate itself. Based on considerations like this, we can see that positive autoregulatory feedback and ultrasensitivity do not, in general, guarantee bistability, though they can provide it.
We have now identified another circuit design principle: Positive, ultrasensitive autoregulation enables bistability.
The toggle switch: a two-gene positive feedback system¶
In natural circuits positive feedback loops are often observed among multiple regulators rather than just a single autoregulatory transcription factor. For example, the “genetic switch” in phage lambda involves reciprocal repression of the cro repressor by the \(\lambda\) repressor, and vice versa (see Mark Ptashne’s classic book, A genetic switch). This arrangement allows the phage to remain in the dormant “lysogenic” state or switch to a “lytic” state in which it replicates and eventually lyses the host cell to infect other cells.
In 2000, Gardner and Collins designed, constructed, and analyzed a synthetic version of this “toggle switch,” and showed that it could similarly exhibit bistability. Here is a simple diagram of the general design.
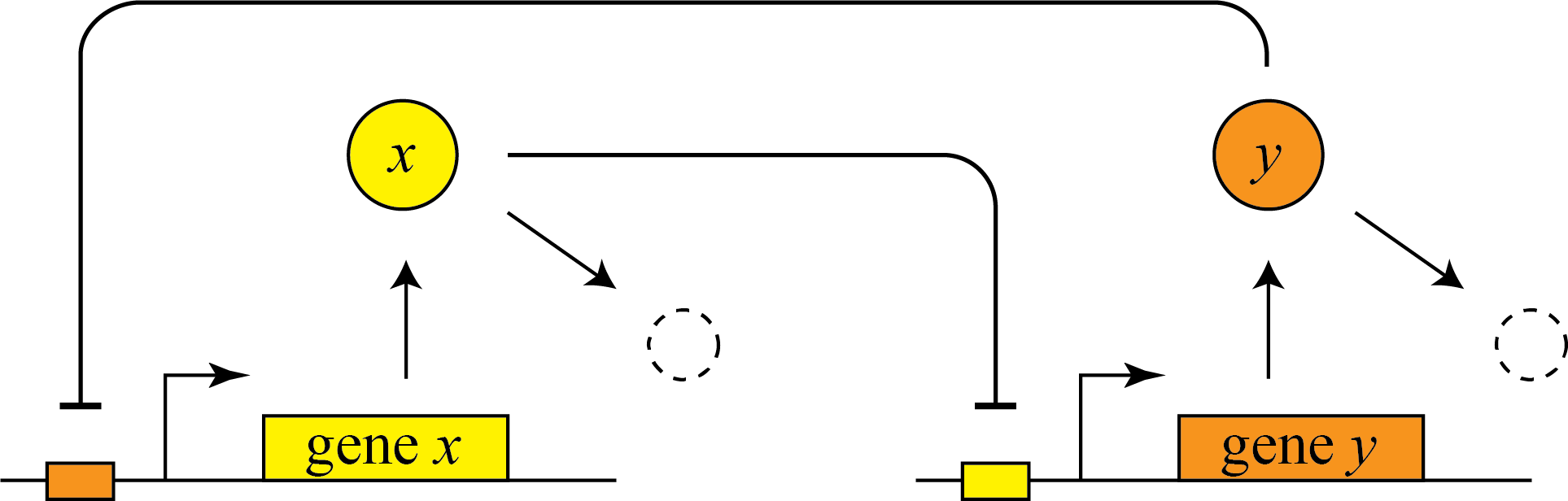
They further showed experimentally that they could put the system into one of two states, by externally activating either of the two proteins. Then they could remove the perturbation and the system stably remained in that state.
To analyze this two-repressor feedback loop, we can write down differential equations for the concentrations of the \(x\) and \(y\) proteins, assuming they have the same Hill coefficient, \(n\).
\begin{align} &\frac{dx}{dt} = \frac{\beta_x}{1 + (y/k_y)^n} - \gamma_x x,\\[1em] &\frac{dy}{dt} = \frac{\beta_y}{1 + (x/k_x)^n} - \gamma_y y. \end{align}
We nondimensionalize by taking \(\beta_x \leftarrow \beta_x/\gamma_x k_x\), \(\beta_y \leftarrow \beta_y/\gamma_y k_y\), \(x \leftarrow x/k_x\), \(y \leftarrow y/k_y\), \(t \leftarrow \gamma_x t\), and \(\gamma = \gamma_y/\gamma_x\). We can now see that the behavior of the system really depends only on (a) the strengths of the two promoters, (b) the relative timescales of the two proteins, and (c) the sensitivity of the response (Hill coefficient):
\begin{align} \frac{dx}{dt} &= \frac{\beta_x}{1 + y^n} - x,\\[1em] \gamma^{-1}\,\frac{dy}{dt} &= \frac{\beta_y}{1 + x^n} - y. \end{align}
A great way to analyze a two-dimensional system like this is by computing the nullclines, which are defined by setting each of the time derivatives equal to zero.
\begin{align} x\text{ nullcline: }& x = \frac{\beta_x}{1 + y^n}, \\[1em] y\text{ nullcline: }& y= \frac{\beta_y}{1 + x^n}. \end{align}
Wherever these two lines cross, one has a fixed point. The stability of that fixed point can be determined by linear stability analysis, which we will introduce in a later chapter. For now, we will plot the two nullclines and investigate their behavior by varying the dimensionless parameters \(\beta_x\) and \(\beta_y\), as well as the Hill coefficient \(n\).
[If we want to have interactivity with a Bokeh plot when we convert the notebook to HTML, we need to use JavaScript callbacks. We do not need to have a Python interpreter running to perform the calculations to update the plot in that case, and the updates are all down by the client’s browser. In the code cell below, we use a CustomJS callback. You can read the code, and the syntax for doing this should be clear.]
[9]:
# Set up sliders
params = [
dict(name="βx", start=0.1, end=20, step=0.1, value=10, long_name="beta_x_slider",),
dict(name="βy", start=0.1, end=20, step=0.1, value=10, long_name="beta_y_slider",),
dict(name="n", start=1, end=10, step=0.1, value=4, long_name="n_slider"),
]
sliders = [
bokeh.models.Slider(
start=param["start"],
end=param["end"],
value=param["value"],
step=param["step"],
title=param["name"],
)
for param in params
]
# Build base plot with starting parameters
beta = 10
n = 4
# Compute nullclines
x_y = np.linspace(0, 20, 400)
y_x = np.linspace(0, 20, 400)
x_x = beta / (1 + y_x ** n)
y_y = beta / (1 + x_y ** n)
source = bokeh.models.ColumnDataSource(data=dict(x_x=x_x, x_y=x_y, y_x=y_x, y_y=y_y))
# Make the plot
p = bokeh.plotting.figure(
frame_height=275,
frame_width=375,
x_axis_label="x",
y_axis_label="y",
x_range=[-1, 20],
y_range=[-1, 20],
)
p.line(x="x_x", y="y_x", source=source, line_width=2, legend_label="x nullcline")
p.line(
x="x_y",
y="y_y",
source=source,
line_width=2,
color="orange",
legend_label="y nullcline",
)
# Callback (uses JavaScript)
js_code = """
// Extract data from source and sliders
var x_x = source.data['x_x'];
var x_y = source.data['x_y'];
var y_x = source.data['y_x'];
var y_y = source.data['y_y'];
var beta_x = beta_x_slider.value;
var beta_y = beta_y_slider.value;
var n = n_slider.value
// Update nullclines
for (var i = 0; i < x_y.length; i++) {
x_x[i] = beta_x / (1 + Math.pow(y_x[i], n));
y_y[i] = beta_y / (1 + Math.pow(x_y[i], n));
}
// Emit changes
source.change.emit();
"""
callback = bokeh.models.CustomJS(args=dict(source=source), code=js_code)
# We use the `js_on_change()` method to call the custom JavaScript code.
for param, slider in zip(params, sliders):
callback.args[param["long_name"]] = slider
slider.js_on_change("value", callback)
bokeh.io.show(bokeh.layouts.column(bokeh.layouts.column(sliders), p))
You can investigate the nullclines by playing with the sliders (even in the HTML rendering of this notebook). For \(\beta_x = \beta_y = 10\) and \(n = 4\), we clearly have three crossings of the nullclines, giving one unstable (the one in the middle) and two stable (the ones on the ends) fixed points.
Play with the \(n\) slider: By moving \(n\) down, you can see that bistability persists at lower Hill coefficients. However, it requires a more delicate balancing act, where the values of \(\beta_x\) and \(\beta_y\) need to be large and close in magnitude. Then, at \(n=1\), bistability is lost completely and the system becomes monostable.
Even at higher Hill coefficients, the strengths of the promoters must still be balanced. If we use widely varying values of \(\beta_x\) and \(\beta_y\), we again have monostability. You can see this by setting \(\beta_x = 10\), \(\beta_y = 1.5\), and \(n = 5\) with the sliders.
Thus, as with the single gene autoregulatory positive feedback loop, ultrasensitivity is necessary but not sufficient for bistability. Later in the course, we will further analyze this system and its stability, learning how to further characterize dynamical systems beyond their nullclines.
Discussion question: What are the advantages of the two-gene toggle switch compared to the single gene positive autoregulation circuit?
Summary¶
Protein degradation and dilution rates determine (and limit) the switching speed of a simple transcriptinally regulated gene.
Design principle: Negative autoregulation accelerates turn-on of a transcription factor.
Design principle: Positive, ultrasensitive autoregulation generates bistability.
Even simple circuits of 1 or 2 genes can generate interesting functional capabilities.
Synthetic circuits can be used to test the functions of simple circuits in living cells.
This is our first foray into the analysis of dynamical systems. As we continue to work with dynamical systems, Strogatz’s book is a great introduction, and includes discussion on using nullclines in the analysis.
Computing environment¶
[10]:
%load_ext watermark
%watermark -v -p numpy,scipy,bokeh,jupyterlab
Python implementation: CPython
Python version : 3.8.8
IPython version : 7.22.0
numpy : 1.19.2
scipy : 1.6.2
bokeh : 2.3.0
jupyterlab: 3.0.11